
Introducció
Un cop acabats amb el metabolisme dels carbohidrats i havent vist també el cicle de Krebs i la fosforilació oxidativa, avui veurem com el metabolisme dels greixos permet introduir intermediaris al cicle que ens permeten obtenir energia, de manera no tant ràpida com el metabolisme de la glucosa però sí en canvi de manera molt més eficient, podent generar molta més energia a llarg plaç.
Lectures prèvies recomanades
- Apunts de bioquímica – Els lípids i els enzims
- Apunts de fisiologia de l’exercici – Introducció al metabolisme energètic
- Apunts de fisiologia de l’exercici – Glicòlisi Aeròbica i el Cicle de Krebs
- Apunts de fisiologia de l’exercici – Fosforilació Oxidativa i la Cadena de Transport d’Electrons
Emmagatzemament i degradat dels greixos
Els greixos representen la reserva d’energia més important de l’organisme en termes quantitatius. Mentre que les reserves de glucogen hepàtiques i musculars poden proporcionar de 1.200 a 2.000 kcal d’energia, el greix emmagatzemat al múscul i en els adipòcits pot arribar a aportar entre 70.000 i 75.000 kcal.
Els greixos són una font d’energia quasi bé inesgotable, però la seva utilitat està condicionada per la localització de les reserves lipídiques.
.jpg) Aquests adipòcits es troben presents als músculs en menor mesura, per la qual cosa l’organisme ha d’articular mecanismes de transport cap al territori muscular actiu, que fan que la dinàmica de la via sigui més lenta.
Aquests adipòcits es troben presents als músculs en menor mesura, per la qual cosa l’organisme ha d’articular mecanismes de transport cap al territori muscular actiu, que fan que la dinàmica de la via sigui més lenta.
 S’emmagatzemen principalment en forma de triglicèrids al teixit adipós i en petites quantitats en d’altres teixits.
S’emmagatzemen principalment en forma de triglicèrids al teixit adipós i en petites quantitats en d’altres teixits.
Les dues principals característiques que diferencien i fan una font ideal d’energia als greixos respecte als carbohidrats pel que fa a esforços sostinguts en el temps son:
- Els lípids contenen més del doble d’energia per unitat de pes que els hidrats de carboni
- Els lípids no s’emmagatzemen hidratats a l’organisme
Els compostos denominats lípids són un grup molt ampli de substàncies insolubles a l’aigua, però sí solubles en diferents dissolvents orgànics (si aquests conceptes no us resulten familiars, no està de més donar una ullada als apunts de bioquímica dels lípids).
Els fonamentals com a substrat energètic són els àcids grassos lliures, que s’emmagatzemen com a triglicèrids.
Els greixos com a substrat no poden actuar amb la urgència que ho fan els carbohidrats. Aquesta realitat provoca que ens trobem davant d’un combustible que respon davant els exercicis que es mantenen durant el temps, però que no poden ajustar-se totalment a esforços més breus e intensos.
D’entre tots els tipus de greixos que existeixen, només els triglicèrids responen a la funció energètica. Aquesta molècula està composada per glicerol i 3 àcids grassos (AG), però en el metabolisme energètic hi participen especialment els àcids grassos.
Per a realitzar això, l’enzim lipasa sensible a les hormones s’encarrega de la lipòlisi (degradació, catabolisme) del triglicèrid, “trencant” la molècula per a que apareguin el glicerol i els 3 àcids grassos que es podran bolcar al torrent sanguini.
Una part del glicerol queda dissolt en plasma i l’altre es dirigirà al fetge per a sotmetre’s al procés de la gluconeogènesi que el convertirà en glucogen. El glicerol, doncs, no és un substrat energètic vàlid per a les fibres musculars, però pot participar en canvi en els processos d’obtenció de glucosa hepàtica.

 D’altra banda, els àcids grassos es combinaran amb l’albúmina (no he tingut l’oportunitat de pujar els apunts d’anatomia sobre això pertinents però si fos el cas sabríem ja que l’albúmina actua com a proteïna transportadora) formant un complex que es trasllada a través de la sang i que anomenem àcids grassos lliures (free fatty acids en anglès i típicament abreujat com FFA).
D’altra banda, els àcids grassos es combinaran amb l’albúmina (no he tingut l’oportunitat de pujar els apunts d’anatomia sobre això pertinents però si fos el cas sabríem ja que l’albúmina actua com a proteïna transportadora) formant un complex que es trasllada a través de la sang i que anomenem àcids grassos lliures (free fatty acids en anglès i típicament abreujat com FFA).
L’albúmina és una proteïna amb una funció molt important doncs atorga als AG la solubilitat necessària per a que a través del plasma puguin arribar als músculs.
Una vegada els AG es troben al múscul són activats enzimàticament amb l’hidròlisi d’ATP. En aquest procés es trenquen dos enllaços fosfat i s’allibera energia en dues ocasions. Com a conseqüència, es produeix AMP (adenosin-monofosfat) i Pi.
A partir d’aquí, l’AG podrà quedar emmagatzemat al múscul en forma de petites gotes lipídiques per a quan es requereixi, o bé pot seguir directament el procés d’oxidació, generant energia.
Metabolisme dels greixos: la beta-oxidació
El metabolisme dels AG es coneix com a metabolisme aeròbic. És un procés que comença quan l’enzim acil-CoA sintetasa transforma els AG en acil-CoA. El acil-CoA és introduït dins els mitocondris a través d’un transportador especialitzat com la carnitina.
 La carnitina és un aminoàcid que es produeix en major part per l’organisme però que també es pot aportar a través de la dieta (i suposo que ho estareu pensant: també de la suplementació).
La carnitina és un aminoàcid que es produeix en major part per l’organisme però que també es pot aportar a través de la dieta (i suposo que ho estareu pensant: també de la suplementació).
La carnitina està localitzada en l’espai intermembranal de la cèl·lula i compta amb uns col·laboradors anomenats acil-carnitina transferasses que permeten que el grup acil-CoA mantingui el seu potencial energètic en el seu camí cap a l’interior de la cèl·lula.
Pprimer una acil-carnitina transferassa converteix el acil-CoA en acil-carnitina per a que travessi la membrana cel·lular, i ja en el seu interior, una altra acil-carnitina transferasa permet alliberar el radical acil de la carnitina i formar acil-CoA.
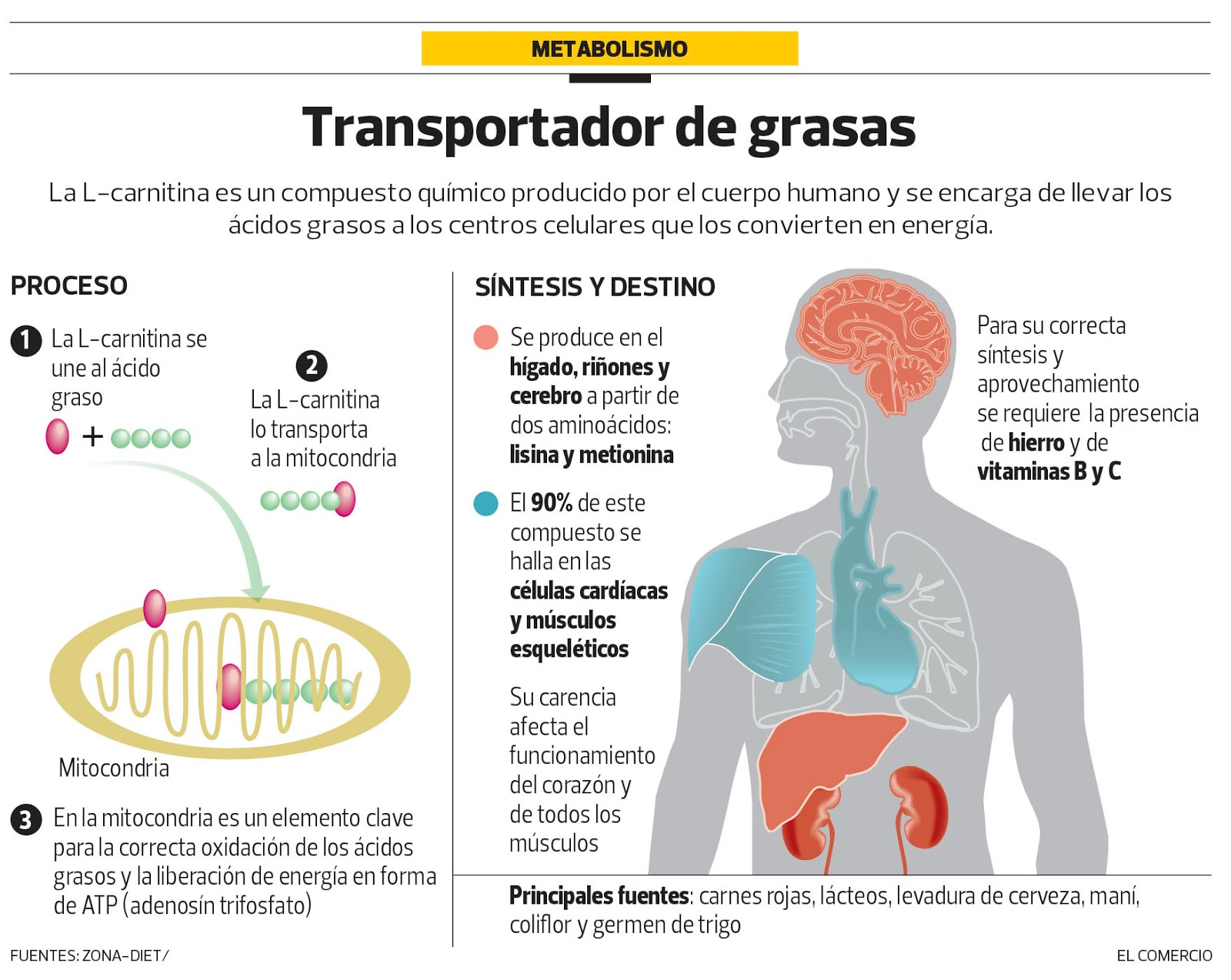
Tot i que aquest mecanisme de difusió no depèn directament de l’oxigen, està vinculat a la capacitat aeròbica del subjecte, degut a que a major volum i densitat mitocondrial, més quantitat d’acil-carnitina transferasses i, per tant, major potencial oxidatiu, ja que es pot utilitzar un major nombre d’àcids grassos per al metabolisme aeròbic.
 Un cop el acil-CoA ha arribat al mitocondri comença el procés anomenat beta-oxidació (representat amb la lletra grega β). Aquesta via metabòlica té caràcter cíclic, doncs el acil-CoA es va oxidant contínuament en el seu carboni β (d’aquí lo de β-oxidació), genera un acil-CoA amb 2 carbonis menys i un acil-CoA.
Un cop el acil-CoA ha arribat al mitocondri comença el procés anomenat beta-oxidació (representat amb la lletra grega β). Aquesta via metabòlica té caràcter cíclic, doncs el acil-CoA es va oxidant contínuament en el seu carboni β (d’aquí lo de β-oxidació), genera un acil-CoA amb 2 carbonis menys i un acil-CoA.
La beta-oxidació es pot aplicar tantes vegades com carbonis es disposi per a reduir. D’aquesta manera, en un AG que tingui 18 àtoms de carboni, es repetirà 8 vegades la β-oxidació i s’aconseguiran per tant 9 acetil-CoA (veure aquest PDF per a descripció més gràfica).
En la β-oxidació del acil-CoA fins a acil-CoA amb dos àtoms de carboni menys i acetil-CoA, s’alliberen hidrògens en dues de les seves reaccions. Això provoca la reducció de un NAD i un FAD i la posterior activació de la cadena de transport d’electrons.
Al final del cicle, el acil-CoA genera un nou procés de β-oxidació i l’acetil-CoA entrarà al cicle de Krebs per a seguir el procés vist en la base aeròbica de la degradació dels carbohidrats. Així doncs, quants més carbonis formin l’àcid gras, més compostos acil-CoA passaran cap a la β-oxidació i més vegades s’activarà el cicle de Krebs.
Per aquest motiu el metabolisme dels greixos està en disposició de generar molta més energia que el de la glucosa.
El balanç d’ATP d’un àcid gras com l’àcid palmític és de 106 ATP aproximadament (+info: càlculs).
Metabolisme dels cossos cetònics
 Els cossos cetònics es formen al fetge com a resultat de la degradació dels àcids grassos lliures i s’entreguen al torrent sanguini per a servir com a font d’energia alternativa al metabolisme de la glucosa quan aquesta és escassa.
Els cossos cetònics es formen al fetge com a resultat de la degradació dels àcids grassos lliures i s’entreguen al torrent sanguini per a servir com a font d’energia alternativa al metabolisme de la glucosa quan aquesta és escassa.
Durant l’exercici de curta durada els cossos cetònics intervenen molt poc en el metabolisme dels músculs exercitants, inclús en altes concentracions en sang, no superant en general el 7% del total d’energia emprada (N. del T: encara està vigent avui dia aquesta afirmació? deures!).
L’entrenament de resistència augmenta el contingut d’enzims emprats en la utilització de les cetones. En subjectes entrenats que realitzen exercici de resistència, l’augment de les cetones en sang no és tant marcat com en els desentrenats, i això és degut al consum i utilització de les mateixes pel múscul esquelètic.
Cal tenir en compte que les concentracions elevades de cossos cetònics durant l’exercici poden ser utilitzades com a font d’energia per d’altres teixits no musculars com el cervell, quan hi ha mancança de glucosa.
Els apunts entren molt poc en el rol del metabolisme dels cossos cetònics en l’exercici ja que suposadament aporten poca energia a l’activitat i el seu ús comporta entrar en un estat de mancança de glucosa que, en general, es procura evitar doncs implicaria pitjor rendiment atlètic.
Alguns investigadors estan barallant l’hipòtesi que portar la contraria a aquesta estratègia i potenciar al màxim el metabolisme dels greixos -i en particular explotar l’ús dels cossos cetònics- a través de l’entrenament i la dieta podria donar millores en el rendiment a llarg plaç quan l’atleta està adaptat, la qual cosa requereix força temps.
A manca d’una entrada d’opinió sobre cetosi nutricional (que encara no m’he atrevit a fer degut a que no tinc una opinió 100% i menys encara totalment fonamentada), podeu curiossejar la secció d’estudis científics que vaig recopilant sobre la matèria.
Deixo una revisió molt recent i força interessant (de lliure accés) que precisament discuteix sobre això (Cox & Clarke, 2014).
Ús dels greixos durant l’exercici
Com hem comentat ja anteriorment, el rendiment energètic d’un lípid és considerable, però perd eficiència per la lentitud associada als seus processos de combustió.
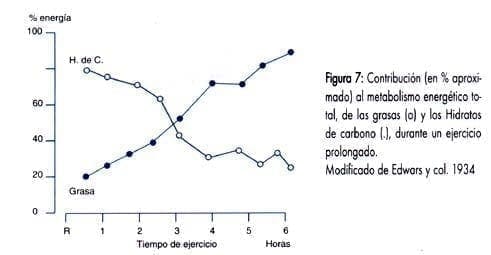
Tot i que la participació d’aquest combustible existeix des de l’inici de l’exercici, la tassa lipolítica no és realment significativa fins que l’esforç es manté durant aproximadament més de 30 minuts.

Imatge agafada del blog dfarmacia.com
La regulació de la via aeròbica depèn també d’elements hormonals. L’activitat lipolítica s’accelera quan s’incrementa la funció simpàtico-adrenal i tendeix a inhibir-se amb la presència d’insulina en sang.
A partir d’aquests marcadors, cada individu s’aprofita de la seva capacitat metabòlica particular, la qual està relacionada amb la capacitat de transport d’àcids grassos per la sang.
Aquesta capacitat depèn, entre d’altres, de la concentració d’albúmina i del flux sanguini en el teixit adipós.

(N. del T.) Entenc que se surt de l’abast dels apunts entrar en això però crec que estaria bé tenir altament present que aquesta descripció encaixa bé en condicions ideals -subjectes “metabòlicament sans”- i que en canvi aquesta capacitat per a utilitzar els greixos es podria veure altament alterada en persones amb obesitat (síndrome metabòlica).
D’alguna manera, els apunts fan un petit incís en lo de “la seva capacitat metabòlica” que per això he subratllat i posat en negreta. En la meva opinió, vincula totalment amb aquest fet. Per a la majoria, representa una capacitat subproducte de les adaptacions de l’entrenament en conjunt amb la genètica de l’individu, però… i l’impacte de la dieta al llarg de la vida?
Un excés d’aliment en conjunt amb una dieta massa alta en carbohidrats sembla ser avui dia un dels motius que condueixen a la SM (Freedland, 2004). Aquí trobem un vincle sobre com això ens podria fer més ineficients utilitzant el metabolisme dels greixos.
Els àcids grassos seran degradats a les fibres ST durant les activitats d’intensitat moderada. Això té importància per a la programació de l’entrenament i també per a la construcció de programes de salut.
Aquestes estratègies no han d’oblidar que en d’altres exercicis més intensos els carbohidrats són els majors responsables d’oferir energia, pel que els intents de pèrdua de pes han de recórrer a exercicis aeròbics, que utilitzin els greixos com a substrat principal (uhm… la qual cosa ens retorna al debat de cardio lent i sostingut vs alta-intensitat, em penso que en Kiefer no estaria d’acord amb això, tot i que en Lyle McDonald ja ha contra-argumentat per què finalment això seria així).
 En el marc del rendiment esportiu, un entrenament de resistència permet a l’esportista la utilització dels lípids com a font d’energia. Això resulta molt interessant, doncs habituem a l’atleta a estalviar glucogen per als esforços d’alta qualitat i per tant estem en disposició de retardar la fatiga.
En el marc del rendiment esportiu, un entrenament de resistència permet a l’esportista la utilització dels lípids com a font d’energia. Això resulta molt interessant, doncs habituem a l’atleta a estalviar glucogen per als esforços d’alta qualitat i per tant estem en disposició de retardar la fatiga.
Aquesta capacitat de preservar els dipòsits de glucogen a partir de la dinamització del metabolisme aeròbic significaria que utilitzant els AG s’inhibeix en cert grau la glucòlisi, la glucogenòlisi i la oxidació del piruvat.
L’acumulació de citrat relacionada amb la degradació dels greixos inhabilita l’enzim PFK, amb la qual cosa s’acumula glucosa-6-fosfat i s’inhibeix a la vegada la hexocinasa per a no degradar més glucosa.
Material extra
Com haureu notat, en aquesta ocasió no m’he pogut estar de introduir alguns enllaços externs per a estendre una mica més els conceptes de la unitat. Sigui com sigui, i per tal de resumir i aclarir els conceptes vistos en aquesta entrada, que forma part de l’interminable unitat dos de l’assignatura, us deixo ja l’habitual recopilació de vídeos externs.
També us deixo un anàlisi interessant amb el que m’he topat sobre la regulació i limitacions de l’oxidació dels AG durant l’exercici (Jeppesen & Kiens, 2012) que bàsicament explica el mateix que aquesta entrada, però en anglès i millor 😉
Pel que fa al tema d’avui, la sèrie de vídeos que hem estat veient en les darreres entrades del Canal Ciència només fan un petit incís a l’ús dels greixos en la segona meitat de la part 6 del documental. Tot i així, no està de més donar-li una ullada per a veure com encaixa tot plegat!
Altres vídeos interessants:
- Bozeman Science – Lipids -> Quasi que vincula més amb els apunts de bioquímica però no va malament, tot va lligat
- The Energy Systems Explained -> Resum de tot plegat junt amb les entrades anteriors
- Formation of Acetyl-CoA by KINprof – Sèrie de 5 vídeos sobre el metabolisme del lactat
- Handwritten Tutorials – Cellular Respiration 4: Beta-Oxidation -> Aquest és el vídeo que realment cobreix millor el resum de l’entrada
- Beta Oxidación de Ácidos Grassos per Dr Luca Merlini -> en castellà
- Beta-Oxidación -> Més de lo mateix
- Beta-Oxidación -> Historia del “ácido grasito” xD
- Universidad de los Andes – Metabolismo de los lípidos -> 1h30 per si no teniu res més que fer… 🙂


















